Article Text

Abstract
Background Aneurysm wall degeneration is linked to growth and rupture. To address the effect of aspirin (ASA) on aneurysm formation under various wall conditions, this issue was analyzed in a novel rabbit bifurcation model.
Methods Bifurcation aneurysms created in 45 New Zealand White rabbits were randomized to vital (n=15), decellularized (n=13), or elastase-degraded (n=17) wall groups; each group was assigned to a study arm with or without ASA. At follow-up 28 days later, aneurysms were evaluated for patency, growth, and wall inflammation at macroscopic and histological levels.
Results 36 rabbits survived to follow-up at the end of the trial. None of the aneurysms had ruptured. Patency was visualized in all aneurysms by intraoperative fluorescence angiography and confirmed in 33 (92%) of 36 aneurysms by MRI/MRA. Aneurysm size was significantly increased in the vital (without ASA) and elastase-degraded (with and without ASA) groups. Aneurysm thrombosis was considered complete in three (50%) of six decellularized aneurysms without ASA by MRI/MRA. Locoregional inflammation of the aneurysm complex was significantly reduced in histological analysis among all groups treated with ASA.
Conclusion ASA intake prevented inflammation of both the periadventitial tissue and aneurysm wall, irrespective of initial wall condition. Although ASA prevented significant growth in aneurysms with vital walls, this preventive effect did not have an important role in elastase-degraded pouches. In possible translation to the clinical situation, ASA might exert a potential preventive effect during early phases of aneurysm formation in patients with healthy vessels but not in those with highly degenerative aneurysm walls.
- aneurysm
- inflammation
- artery
- blood flow
- magnetic resonance angiography
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Aneurysmal subarachnoid hemorrhage due to rupture of an intracranial aneurysm remains one of the most devastating neurological diseases with a high associated mortality and morbidity.1 2 Chronic inflammatory processes of the vessel wall are known to have an important role in intracranial aneurysm wall pathobiology that leads to further growth and rupture.3 4 In this context, the role of aspirin (ASA) as a potential protectant against intracranial aneurysm rupture by antagonizing the inflammatory microenvironment has been elucidated in previous animal studies, human case–control studies, and histological analyses of human aneurysm domes sampled from unruptured and ruptured intracranial aneurysms.3 5–8
For instance, continuous ASA intake for 12 weeks was shown to reduce macrophage-mediated aneurysm wall signal intensity in MRI of human unruptured intracranial aneurysms.9 Additionally, compared with unruptured aneurysms, upregulation of prostaglandin E2 synthase-1 (PGES-1) and cyclooxygenase-2 (COX-2) was observed in ruptured specimens.5 Compared with the control group, regular ASA intake 3 months before microsurgical clipping decreased vessel wall signal intensity on MRI, lowered PGES-1 and COX-2 concentrations, and decreased levels of macrophages in aneurysm tissues.10 In a study of patients taking ASA for 6 months or more, similar vessel wall results were noted using high-resolution 3 Tesla (T) MRI.11 A 10-year retrospective review of patients with multiple intracranial aneurysms showed that ASA significantly decreased the aneurysm growth rate over time.12
Nonetheless, information is lacking about the effect of ASA intake on the natural course of bifurcation aneurysms with vital and already degraded wall conditions. Therefore, in this experimental bifurcation aneurysm rabbit model, we specifically analyzed inflammation and aneurysm growth over time in groups with varied wall characteristics.
Materials and methods
Study design
To analyze the effects of ASA on aneurysm formation under various wall conditions in a rabbit model, bifurcation aneurysms were created in 45 New Zealand White rabbits (figure 1). Animals were randomly allocated to one of three groups as vital (n=15), decellularized (n=13), or elastase-degraded (n=17) using a web-based randomization system (www.sealedenvelope.com). Each group was then assigned to a study arm with or without ASA (n=6).
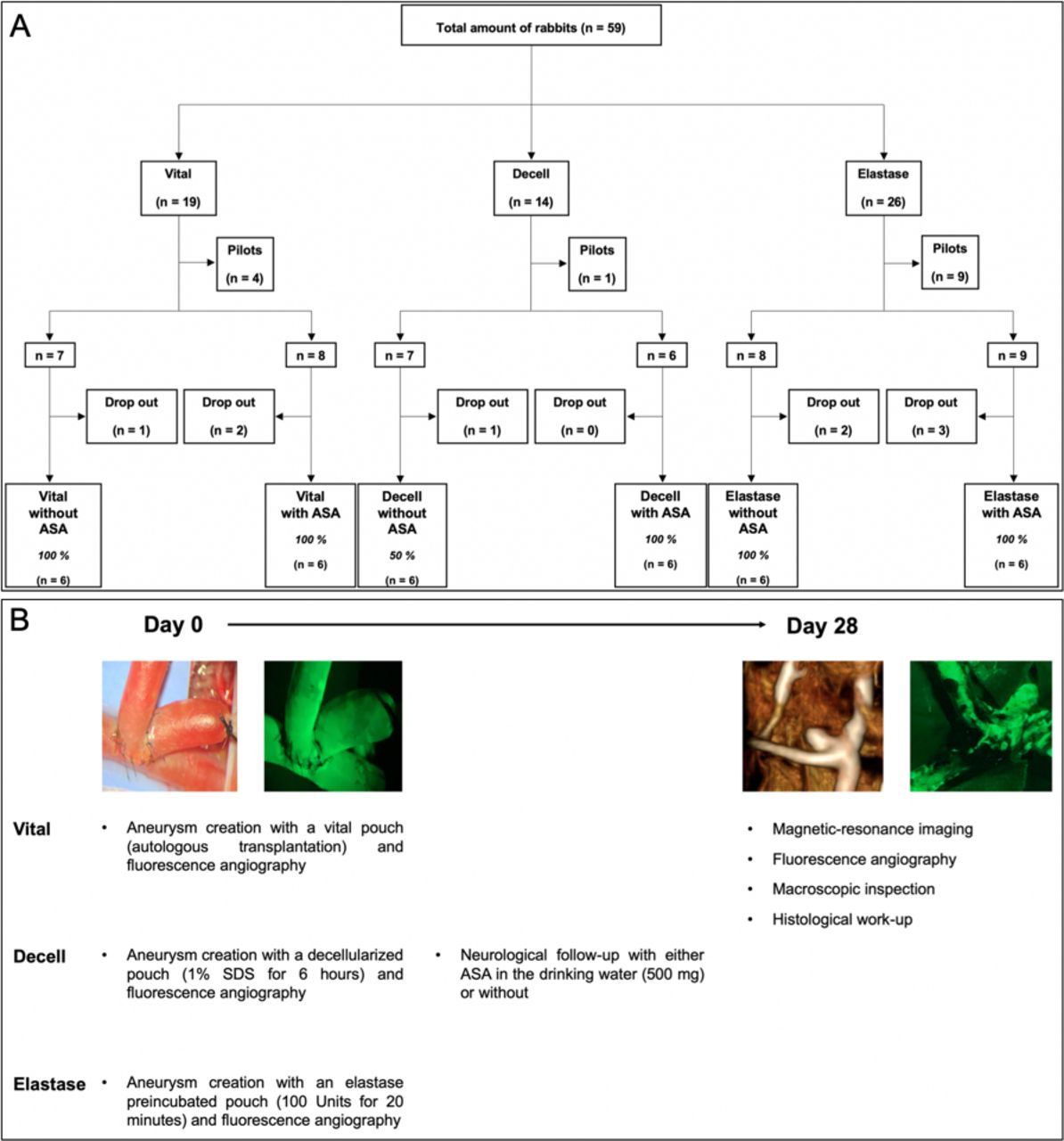
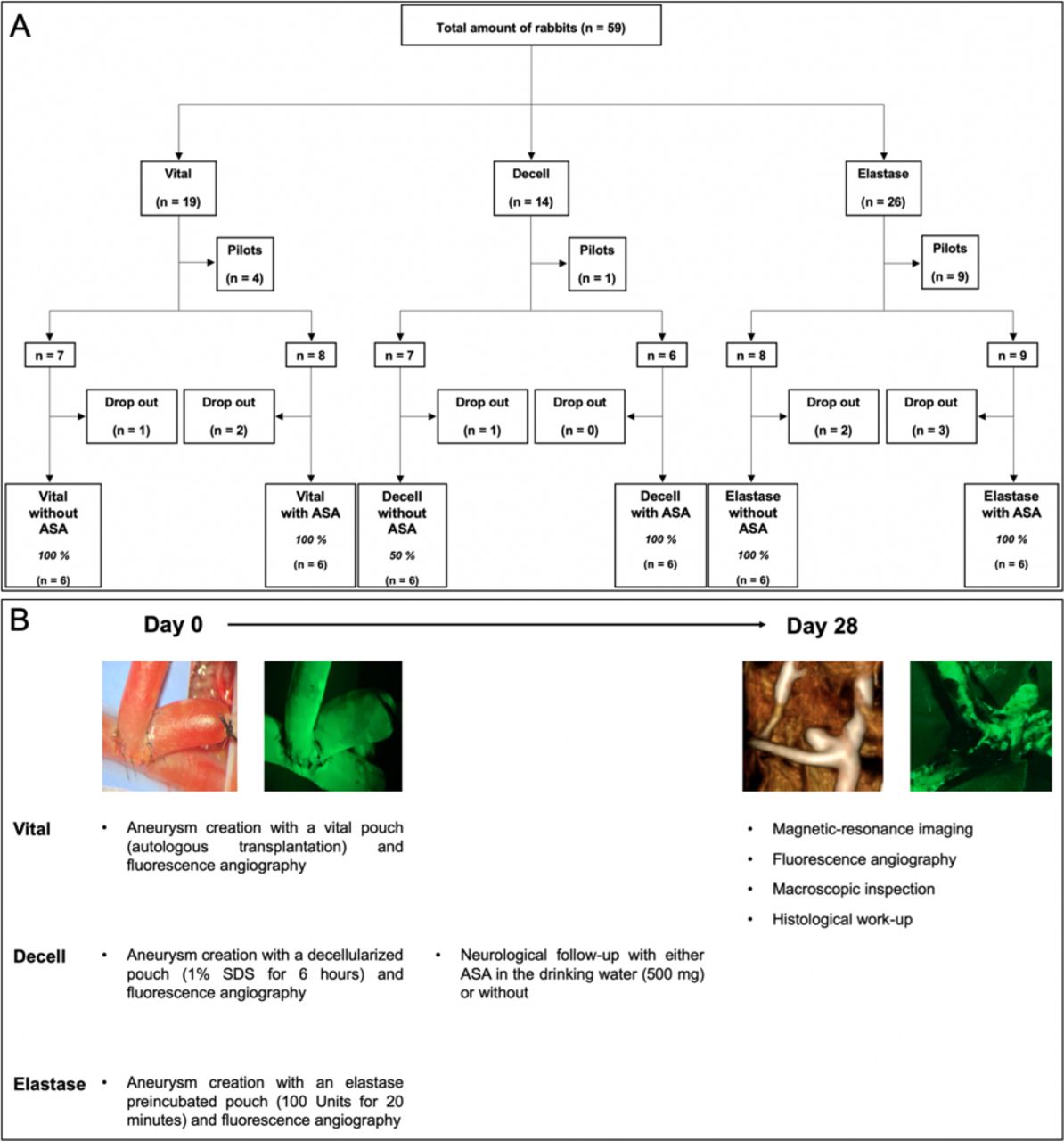
. Flowchart of the study design and follow-up modalities. Of 59 total animals, 36 were included for final analysis and 23 were excluded (ie, 14 in pilot study and 9 early dropouts (2 with complete thrombosis of the left parent artery; 3 with neurologic worsening; 1 with intraoperative digestion of the left carotid artery with subsequent euthanasia; 3 deaths of unknown causes) (Panel A). Patency of the aneurysm complex was confirmed via fluorescence angiography at day 0 intraoperatively, and with magnetic resonance angiography and fluorescence angiography 28 days after surgery (Panel B).
Primary outcomes were defined as aneurysm patency and aneurysm growth 28 days after its creation based on MRI findings. Secondary outcomes were aneurysm inflammation and thrombus organization. Patency of the aneurysm and its parent artery complex was controlled by fluorescence angiography at intraoperative creation and assessed at follow-up after terminal MRI.13 The study design is shown in figure 1.
Experiments were approved by the Local Committee for Animal Care of the Canton Bern, Switzerland (BE 108/16). The 45 female rabbits with a mean weight of 3722.64 g (±276.34 g) and mean age of 16 weeks (±3 days) were housed at room temperature of 22–24°C with a 12-hour light/dark cycle. The rabbits had free access to a pellet and hay diet as well as water. All animals received daily care in accordance with the local institutional guidelines. Animals allocated to the ASA groups exclusively had access to ASA water. Therefore, correct intake of ASA was calculated by monitoring drinking water twice daily. Surgeries were conducted in the Experimental Surgery Facility of the same department under supervision of a board-certified veterinary anesthesiologist. The ARRIVE guidelines were strictly followed.14
Bifurcation aneurysm model and surgical technique
Intraoperative surgical characteristics were recorded (see online supplemental figure 1). Detailed techniques of graft harvesting, graft preparation, and microsurgical creation of the arterial bifurcation have been described elsewhere.15 Briefly, bifurcation aneurysms were created by end-to-side anastomosis of the right common carotid artery (CCA) to the left CCA, and interposition of an arterial pouch in 36 female New Zealand White rabbits (see online supplemental figure 2 and online supplemental video 1).
Supplemental material
Supplementary video
Arterial wall degradation protocols
Considering the protocol without prior treatment, the autologous graft was harvested and directly transplanted to the same animal (autograft). Treatment followed either the sodium dodecyl sulfate (SDS) or the elastase protocol. For the SDS administration, tissue was decellularized according to a previously published protocol.4 For elastase administration, graft decellularization followed the details of a previously published protocol as well (detailed protocol in Supplementary Materials).15
Magnetic resonance imaging
Before final MRI, all animals were weighed and received a subcutaneous application of 0.1 mg/kg acepromazine (Prequillan, 10 mg/mL, Fatro, Italy) and buprenorphine (Temgesic, 0.3 mg/mL, Indivior AG, Switzerland) (0.03 mg/kg). Scanning time performed via 3 T Magnetom Skyra (Siemens, Munich, Germany) (see detailed protocol in Supplementary Materials) was 45 min per animal. On completion of fluorescence video angiography, the animals were euthanized by intravenous injection of 150–300 mg/kg pentobarbital (Esconarkon, 300 mg/mL, Streuli Pharma, Switzerland).
Measurement of the pre- and post-mortem aneurysm volumes
Aneurysms were measured and photographically documented before implantation. Aneurysm volume was calculated based on the formula: π × height × width/2 × length/2. At final MRI, aneurysm volumes were quantified by 3D reconstruction (VitreaCore, Canon, Tokia, Japan); partial thrombogenesis was detected by this imaging and macroscopic evaluation (see online supplemental figure 3). Post-mortem macroscopical analysis and documentation of the aneurysm volume was carried out to assure a certain comparability.
Histology
For histological processing, paraffin-embedded aneurysms were cut in the axis of the underlying parent artery in 2 μm slices and stained, including hematoxylin-eosin (HE), smooth muscle actin (SMA), Masson-Goldner trichrome (MASA), and von Willebrand factor (F8). The slides were then digitized (Omnyx VL 120, GE, USA) and evaluated with a JVS viewer (JVS view 1.2 full version, http://jvsmicroscope.uta.fi/software/, Finland). The light microscopy findings were analyzed qualitatively by two observers (SW and SS) and rated with a previously used four-tier grading system.4 However, our study design precluded blinding of the three groups and the effect of ASA. Periadventitial inflammation, aneurysm wall inflammation, and neutrophils in the thrombus were evaluated in HE-stained slices, and periadventitial fibrosis was evaluated in MASA-stained slices. The presence of hematoma, cellularity, and dissection of the aneurysm wall and luminal thrombus were analyzed in HE-stained slices, whereas endothelium and neointima formation were evaluated in MASA-stained slices. Finally, aneurysm wall cellularity was assessed in SMA-stained slices and endothelium in F8-stained slices.
Statistical analyses
All statistical analyses were calculated using the non-parametric Wilcoxon–Mann–Whitney U test and/or parametric unifactorial variance analysis. A p value of <0.05 and <0.01 was considered significant. Data were analyzed by IBM SPSS Version 22 (USA). The sample size per group was determined using an a priori sample size calculation (BiAS.for.Windows Version 11, Germany). To achieve α=0.05 at β=0.2 with a sigma of 0.2, the sample size calculation showed that 4–8 animals per group were appropriate to achieve a delta between 0.3 and 0.5. Figures were visualized by Graph Pad Prism 8, Version 8.2.0.435 (GraphPad software, USA). All values are expressed as mean±SD or median (IQR).
Results
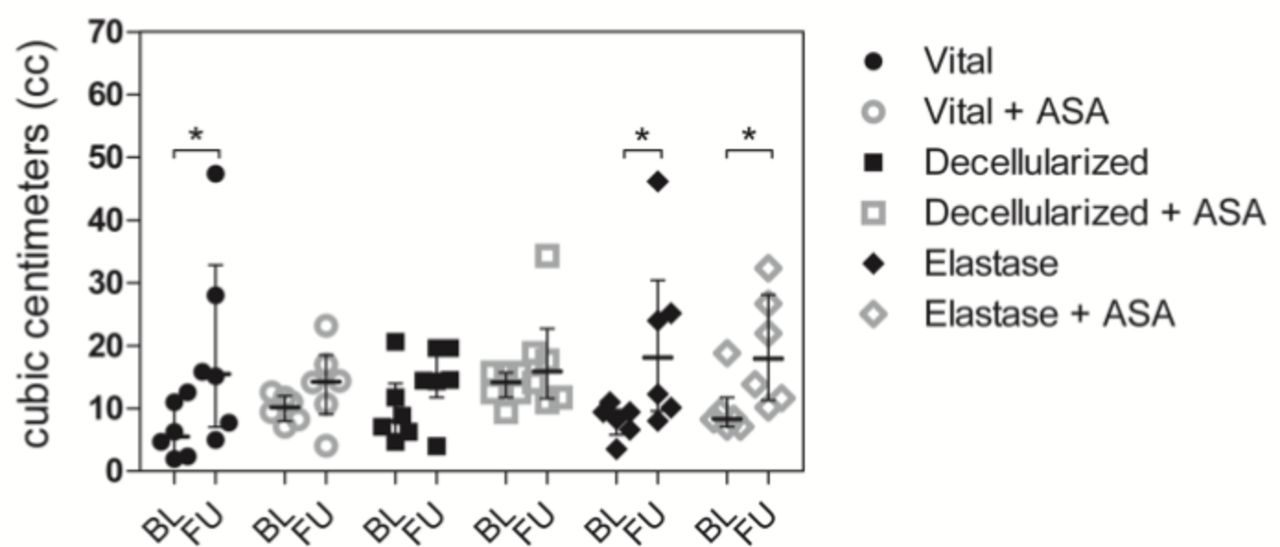
Aneurysms in the vital group without ASA as well as in the elastase groups without and with ASA showed significant growth patterns over 28 days (figure 2). This effect was not observed in the vital group with ASA, nor in the decellularized groups without and with ASA.
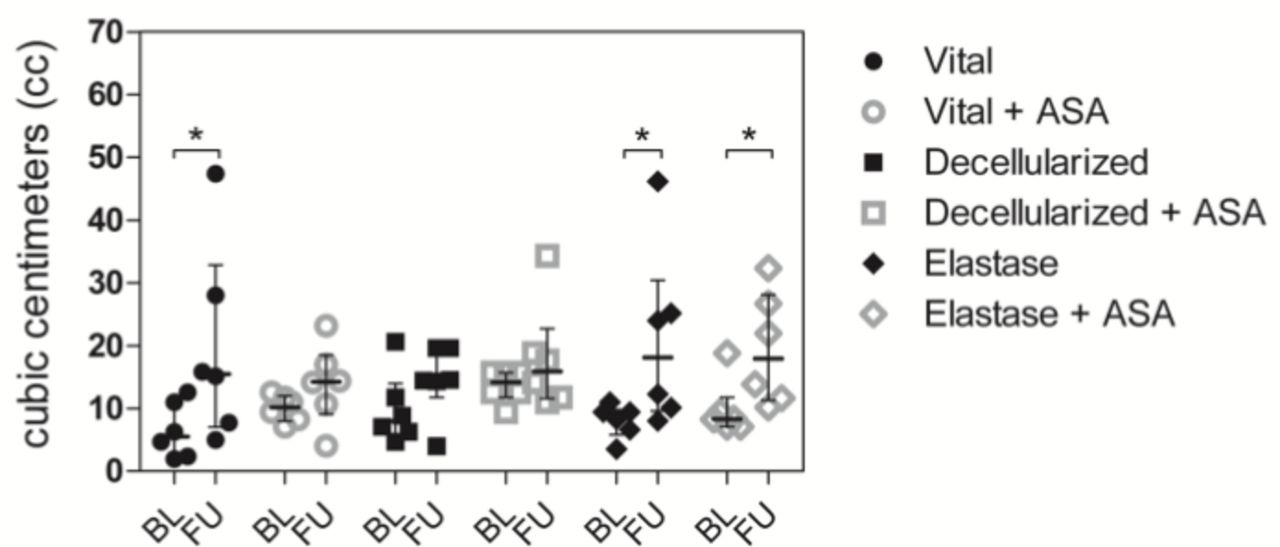
. Bar graph comparing aneurysm volume (cc) at baseline (BL) and follow-up (FU). Vital aneurysms without ASA treatment showed significant patterns of growth whereas those with ASA treatment did not grow. All aneurysms with SDS-degenerated walls (with and without ASA treatment) did not grow. By contrast, all aneurysms that were degraded with elastase (without and with ASA treatment) showed significant growth during follow-up. All values are given as cubic centimeters (cc) and presented as median and interquartile range. * p < 0.05 vs. corresponding group.
Follow-up macroscopic measurements
Aneurysm size increased in all study groups from creation to follow-up (see online supplemental table 1). Specifically, aneurysm size increased in the vital group without ASA from 6.48±4.43 mm3 at creation to 19.85±15.69 mm3 at follow-up (p<0.05) and with ASA from 10.01±2.13 mm3 at creation to 13.91±6.38 mm3 at follow-up (p=0.149). Aneurysm size increased in the elastase group without ASA from 8.03±2.64 mm3 at creation to 20.95±14.31 mm3 at 28-day follow-up (p<0.05) and with ASA from 9.91±4.48 mm3 at creation to 19.46±8.99 mm3 at follow-up (p<0.05). Aneurysm size increased, although not significantly, in the decellularized group without ASA from 9.88±5.79 mm3 at creation to 14.43±9.02 mm3 at follow-up (p=0.261) and with ASA from 13.61±2.56 mm3 at creation to 17.96±8.61 mm3 at follow-up (p=0.41) (see online supplemental table 1).
After 28 days, 100% patency was observed in the vital and elastase groups with or without ASA and in the decellularized group with ASA. Complete thrombosis of the aneurysm and the parent artery was observed in three (50%) of six animals in the decellularized group without ASA. Partial thrombosis was observed in three of six aneurysms in the decellularized group without ASA, in one (16%) of six aneurysms in both the vital group with ASA and the elastase group without ASA, and in all six aneurysms in the decellularized group with ASA. Complete and partial thrombosis of the aneurysm was visualized by contrast-enhanced MRI followed by fluorescence angiography.
Follow-up histological findings
Histological grading included periadventitial inflammation, aneurysm wall inflammation, neutrophils in the thrombus, and periadventitial fibrosis (figure 3).

. Bar graph depicting histological grading for periadventitial inflammation, aneurysm wall inflammation, neutrophils in the thrombus and periadventitial fibrosis. Analyses are grouped for vital, decellularized, and elastase-treated aneurysms without and with ASA treatment using a previously described 4-point grading system (0 = none, 1 = mild, 2 = moderate, 3 = severe). * p < 0.05, ** p < 0.01 vs. corresponding group. (A) Periadventitial inflammation. ASA intake significantly reduced periadventitial inflammation in the vital, decellularized, and elastase groups. (B) Aneurysm wall inflammation. ASA intake significantly reduced aneurysm wall inflammation in the vital, decellularized, and elastase groups. (C) Neutrophils in the thrombus. ASA intake did not significantly reduce neutrophils in the thrombus in the vital, decellularized, or elastase groups. However, a pronounced thrombus invasion of neutrophils was observed in the decellularized group compared with vital and elastase groups. (D) Periadventitial fibrosis. ASA intake significantly reduced periadventitial fibrosis in the vital and elastase groups but not in the decellularized group.
ASA significantly reduced periadventitial inflammation in the vital, decellularized, and elastase groups (p<0.01) (figures 3 and 4). Likewise, ASA also significantly reduced aneurysm wall inflammation in the vital (p<0.01), decellularized (p<0.05), and elastase (p<0.01) groups (figures 3 and 4). However, ASA did not significantly reduce neutrophils in the thrombus in the vital, decellularized, or elastase groups. Invasion of neutrophils was pronounced in the decellularized group compared with the vital and elastase groups (figure 3). Periadventitial fibrosis was significantly reduced in the vital (p<0.01) and elastase groups (p<0.05) and was more pronounced in the vital group (figure 3).

{kind=link}
{kind=link}
{kind=link}
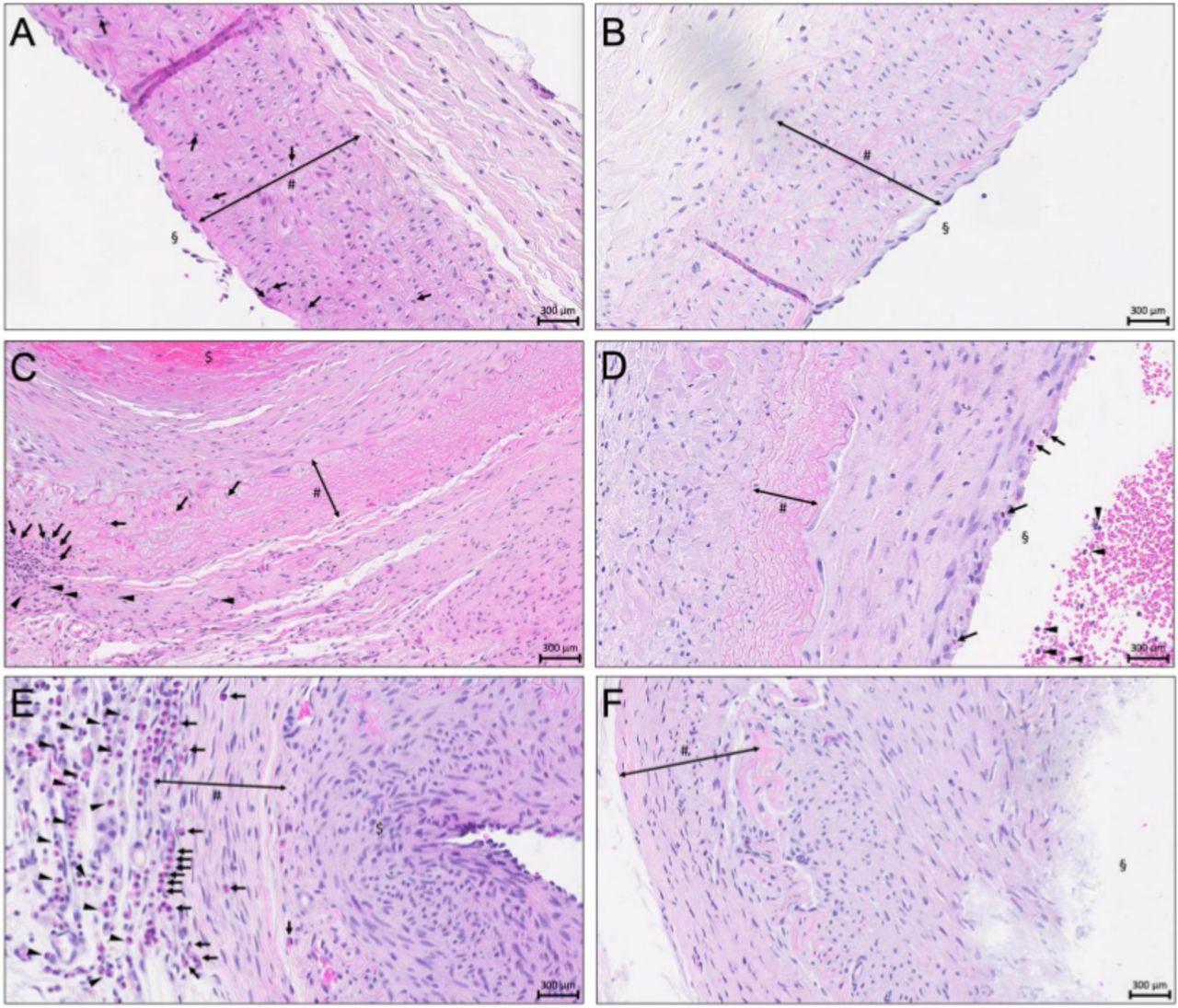{kind=link}
. Representative microphotographs of vital, decellularized, and elastase-treated aneurysms without and with ASA treatment. Hematoxylin-eosin (HE) staining of the media (#). Scale bar: 300 mm.(A) Vital aneurysm without ASA treatment. Aneurysm lumen (§), infiltration of granulocytes (Black arrows).(B) Vital aneurysm with ASA treatment shows no sign of wall inflammation or periadventitial inflammation. Aneurysm lumen (§).(C) Decellularized aneurysm without ASA treatment. Note the apical thrombus formation ($), and intense inflammation of aneurysm wall and periadventitia characterized by accumulating granulocytes (b lack arrows). Periadventitial tissue layer (black arrowheads).(D) Decellularized aneurysm with ASA pretreatment shows no signs of inflammation of aneurysm wall (#) or periadventitia. Note luminal thrombus formation and sparse neutrophils in the outer luminal layer of the thrombus (black arrows) and intraluminally (black arrowheads). Aneurysm lumen (§).(E) Elastase-treated aneurysm without ASA. Beginning luminal thrombus formation ($) with excessive infiltration of neutrophil granulocytes in the aneurysm wall (black arrows and highly degenerated extracellular matrix). Note excessive periadventitial inflammation (black arrow heads).(F) Elastase-treated aneurysm with ASA. Note the highly degenerated matrix in the aneurysm wall. Note absence of inflammation in aneurysm wall and periadventitial. Lumen of the aneurysm (§).
Among grading for other histological characteristics (online supplemental figure 4), we noted that, in all six groups, neither hematoma nor dissection affected the aneurysm wall, endothelial cellularity was not significantly altered, and no significant differences developed in neointima formation. Luminal thrombosis was more pronounced in both decellularized groups. In the vital and elastase groups, neointima formation could not be detected compared with the decellularized groups in which a stronger tendency to neointima formation was noted with ASA (see online supplemental figures 5-7 for illustrative histological samples).
General data and surgical characteristics
Of a total of 45 animals used in the experiments, 14 (4 vital, 1 decellularized, 9 elastase) were used in a pilot study to establish reliable and consistent reproducible procedures. Of 36 animals included in the final study, no deaths occurred intraoperatively and no ischemic brain lesions were detected by MRI in any rabbit before euthanasia.
Among nine (20%) animals that died prematurely and were excluded from analysis, two had complete thrombosis of the left parent artery on postoperative days 3 and 7; three had neurologic worsening and were euthanized postoperatively on days 4, 5, and 8; one had intraoperative digestion of the left carotid artery and was subsequently euthanized; and three died in their cages of unknown causes postoperatively on days 2, 7, and 11.
Intraoperatively, 10 (23%) of 44 aneurysms showed no contrast enhancement in the arterial pouch on fluorescence angiography. These 10 aneurysms then underwent reclamping, reopening, and flushing with heparinized saline followed by thrombus evacuation. Repeat fluorescence video angiography confirmed patency in all 44 aneurysm cases. Physiological values and amplitude of blood pressure were measured in the auricular artery after reopening all vascular clamps.
Follow-up MRI patency rate
After 28 days, 33 (92%) of 36 of aneurysms remained open, two (6%) had complete thrombosis of the aneurysm and its parent vessel complex, and one had complete aneurysm thrombosis. Partial aneurysm thrombosis was observed in 11 (31%) aneurysms, especially among nine (25%) in the decellularized group (online supplemental table 1).
Discussion
This experimental study studied the effects of ASA on bifurcation aneurysms in a rabbit model, specifically the natural course of inflammation and aneurysm growth in groups defined by wall characteristics as vital, decellularized, or elastase degraded. Our results of continuous ASA intake found significantly decreased aneurysm growth in vital aneurysms, mimicking early aneurysmal disease, as well as decreased inflammation in the periadventitia and aneurysm wall irrespective of any of the three initial wall conditions. Interestingly, ASA intake did not attenuate aneurysm growth in elastase-pretreated pouches. The protective effect of ASA in the early stage of aneurysm might be lost in progressive aneurysmal disease.
Vital aneurysms
To analyze potential effects of ASA in the early phase of aneurysm formation, vital arterial pouches were implanted in an artificial arterial bifurcation. Compared with the control group, ASA significantly prevented aneurysm growth and, in histological studies, significantly reduced aneurysm wall inflammation, periadventitial inflammation, and fibrosis.
The potential role of ASA in preventing intracranial aneurysm growth and rupture is multifold.6 First, it acts to block COX-2, which is associated with intracranial aneurysm formation and rupture. Hasan et al showed upregulation of PGES-1 and COX-2 in histological specimens of human ruptured intracranial aneurysms compared with unruptured intracranial aneurysms.5 In tissue samples of patients who underwent intracranial aneurysm clipping, attenuated expressions of PGES-1, COX-2, and macrophages were detected in the wall after 3 months of ASA therapy compared with the control group without ASA. As shown by novel imaging protocols, aneurysm wall inflammation was reduced in patients who received continuous ASA treatment prior to elective clipping.10 With regard to these issues, a vessel wall oriented therapy with ASA might provide an interesting future approach in identifying patients at high risk for intracranial aneurysm rupture and/or assessing treatment response.
Decellularized aneurysms
Decellularized aneurysms did not show significant enlargement with or without ASA treatment. Compared with the control group, inflammation in the aneurysm wall and periadventitia was significantly reduced in the ASA arm. The role and importance of degenerated aneurysm walls in growth and rupture is undisputed.4 16 In general, fewer mural cells implicate a worse biologic response in aneurysm healing because of the inability of intraluminal thrombus to organize. Previously, loss of mural cells was shown to have an important role in furthering its progression in a saccular rat sidewall model with untreated aneurysms,4 after coil embolization,17 and also after stent treatment.18 With decreased numbers, there are fewer cells that can migrate into the thrombus and form a neointima. Furthermore, in a sidewall rabbit model, all aneurysms thrombosed spontaneously.19 In our bifurcation series, in the ASA arm all aneurysms had partial thrombosis whereas the group without ASA had complete thrombosis in half of the aneurysms and partial thrombosis in the other half. Graft decellularization by SDS, including endothelial cells, may have contributed substantially to this high rate of thrombosis. Therefore, other degradative compounds such as elastase pretreatment were further analyzed for a potential ASA effect on aneurysm formation in advanced disease.
Elastase-degraded aneurysms
Compared with vital aneurysms, a protective effect in prevention of aneurysm growth was not observed in elastase-degraded pouches and ASA intake clearly failed to prevent aneurysm growth. After 28 days, significant growth was observed both with and without ASA. Therefore, while ASA intake might be protective during early stages of aneurysm formation with relatively healthy vessel walls, this protective effect is likely to be lost in advanced disease. Nevertheless, aneurysm wall inflammation, periadventitial inflammation, and fibrosis were significantly reduced in the ASA group. This fact suggests that, along with inflammatory components, other factors driving aneurysm enlargement (eg, hemodynamics of a true vessel bifurcation) must also be considered.
In our pilot study using 14 animals, the group of nine elastase-treated animals was the largest; this group also included 5/9 animals that died prematurely and contributed to the overall 15.3% mortality rate. The reasons for the increased mortality, even when performing topical elastase incubation in our model, might be the non-specific effects of elastase in circumferential digestion of all tissues (shown in online supplemental figure 2C), leading to possible thrombogenic reactions at a molecular level.20
Use of ASA in aneurysm treatment and further clinical implications
Several studies have previously shown substantial increases in aneurysm size in deteriorated aneurysm walls in rabbits over time when using adjuncts such as Eastman 910 or elastase after in vitro collagenase I infusion.21 In our bifurcation series, these observations were confirmed in both vital and elastase-degraded pouches. The significant aneurysm growth over time observed in the vital group without ASA treatment was similar to the natural course of bifurcation aneurysms observed in humans.2 Here, the significant upregulation of neutrophils in the periadventitia and aneurysm wall might be the cause. Accordingly, with its anti-inflammatory effects, ASA pretreatment in the vital group thus prevented significant aneurysm growth after 28 days. Aligned with the results of other studies, we consider that, through antagonizing inflammatory processes such as COX-2 and PGES-1 inhibition, a certain degree of aneurysm wall stabilization might be achieved.
In a retrospective review of 146 patients, Zanaty et al observed that ASA intake significantly decreased the rate of intracranial aneurysm growth over time.12 In a large retrospective single-center study of 4701 patients, Can et al found that ASA intake was associated with a decreased risk of intracranial aneurysm rupture.22 Furthermore, Hudson et al clearly demonstrated that ASA intake beneficially modulated the inflammatory microenvironment within the intracranial aneurysm vessel wall, thereby preventing rupture and aneurysm growth in patients with multiple intracranial aneurysms over time.8 These findings were confirmed in other preclinical and clinical investigations.23 24
Comparable with our results, preclinical and clinical studies have reported similar encouraging results when targeting inflammatory processes in the aneurysm wall, especially with ASA, to avoid further growth and rupture; they also hypothesized that its anti-inflammatory effect might be protective in providing aneurysm stability.3 10 More recently, these data were further confirmed in population-based studies of more than 200 000 patients with intracranial aneurysms who received low-dose long-term ASA therapy. Regarding the entire study population, the incidence of subarachnoid hemorrhage by aneurysm rupture was reduced by more than 20%.25 Nonetheless, the benefit of this conservative treatment approach must be confirmed in prospective randomized multicenter trials such as the PROTECT-U trial, which considered the influences of blood pressure reduction in combination with ASA.1
Study limitations
To the best of our knowledge, this study is the first report of a true bifurcation aneurysm model using arterial grafts that allow for a direct comparison of different wall characteristics (vital, decellularized, elastase degraded). Strengths of this model included its relatively large sample size and a multimodal data analysis approach (ie, baseline and follow-up fluorescence angiography, MRI, histological assessment) that provided consistent and reliable values for results and interpretation.
The aim of these experiments was to study a biological concept of vessel wall biology, therefore the results of this extracranial aneurysm model should be extrapolated with caution to a true intracranial aneurysm model. Compared with endovascular approaches for aneurysm induction, surgical dissection causes locoregional inflammation so a surgically-created aneurysm likely causes stronger long-lasting inflammation. To reduce this bias in analyzing the data as far as possible, the final follow-up timepoint was 28 days after surgical creation of the aneurysm. Furthermore, postoperative administration of meloxicam for analgesia may have influenced locoregional inflammation in a beneficial fashion. However, this bias was minimized because all the animals received meloxicam for only 3 days. Regarding the 20% mortality and initial 23% rate of intraoperative thromboses verified by fluorescence angiography, other factors to consider include potential thrombogenic properties of the suture material and modified arterial pouches, and the learning curve required for this sophisticated surgical technique. Of note, initial MRI directly after implanting the arterial pouch would provide the most precise aneurysm volume, but partial thrombosis of the arterial pouch within 28 days detected by terminal MRI might potentially not depict true aneurysm growth over time and could even interfere in the comparison between immediate postoperative MRI and MRI performed at terminal follow-up. Lastly, immediate postoperative MRI was not performable because of missing availability, so exact macroscopic measurement was performed every time prior to implantation and also after terminal MRI and explantation of the aneurysm complex to detect macroscopic pattern of growth. Therefore, a certain reproducibility should be given but, nevertheless, macroscopic measurement after pouch implantation intraoperatively as well as postoperatively after 28 days would eventually be more precise.
Besides the use of neutrophils to monitor the indirect effects of ASA on the inflammatory microenvironment, many specific parameters (eg, COX-1, COX-2) can now be measured to identify direct effects. Nevertheless, previous studies have identified neutrophils as the driving factors for aneurysm growth. Potential molecular targets of ASA in cerebral aneurysm pathophysiology are manifold. For example, prostaglandin E2 targeted by ASA increases vascular permeability and attracts/activates both macrophages and neutrophils. Therefore, an indirect effect of ASA intake should be at least depicted.
Finally, although we aimed for maximal standardization, we cannot exclude the possibility that slight differences in aneurysm volumes might have been hemodynamically relevant by causing different shear stress.
Conclusions
We believe this to be the first report of a true bifurcation aneurysm model using arterial grafts that allowed for a direct comparison of different wall characteristics (vital, decellularized, elastase-degraded). Continuous ASA administration reduced inflammation of the aneurysm wall irrespective of the initial wall condition in a rabbit true bifurcation aneurysm model. ASA significantly prevented aneurysm growth in aneurysms with vital walls, whereas this preventive effect did not seem to play an important role in elastase-degraded pouches. Considering these findings, the potential preventive effect of ASA in the early phase of aneurysm formation in patients with healthy vessels may be lost in those with highly degenerative aneurysm walls.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
Ethics statements
Patient consent for publication
Ethics approval
The project has been performed according to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and was performed in accordance with the National Institutes of Health Guidelines for the care and use of experimental animals and with the approval of the Animal Care Committee of the Canton Bern, Switzerland (Approval No. BE 108/16).
Acknowledgments
We are deeply grateful to: Olgica Beslac and Kay Nettelbeck for their excellent support and technical assistance during the peri-operative phase; Alessandra Bergadano, DVM, PhD, for the dedicated supervision of the long-term animal health and Mary Kemper for editing and proofreading.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors Conception and design: SW, SM. Experimental procedures: SW, SM, BEG, FS. Histological sample preparation and analysis: SW, HRW, MvG, GB, LA, SS. Drafting the article: SW, BEG, SM. Statistical analysis and interpretation of data: SW, BEG, LA, SM. Critically revising the article: JF, HRW, SM, LR, BEG, JR, DC. Administrative support: SM.
Funding This work was supported by the Swiss National Science Foundation SNF (310030_182450). The authors are solely responsible for the design and conduct of the presented study and declare no competing interests.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.