Article Text

Abstract
Background The extent, rate, and source of endothelialization following coil embolization of saccular aneurysms remains poorly understood. We performed a whole tissue mount, dual immunohistochemical analysis of experimental aneurysms to characterize the state of endothelialization over time after platinum coil embolization.
Method and material Elastase-induced rabbit aneurysms were created and treated with bare platinum coils. Samples were harvested at 4 and 8 weeks (n=6 for each). En face whole tissue mount staining with antibodies to CD31 and α-smooth muscle actin was used to identify endothelial cells and smooth muscle cells, respectively. Sytox green stain was used to demonstrate nuclear morphology for identification of inflammatory cells. The extent of endothelialization was measured in relation to the aneurysm neck–parent artery interface.
Results At 4 weeks after coil embolization, very localized membranous tissue and neoendothelial cells were detected on the coil loops immediately adjacent to the parent artery–neck interface, but the remainder of the coil loops remained devoid of endothelial cells. At 8 weeks neoendothelial cells were more confluent over the coils than at 4 weeks, and extended up to 900 µm from the parent artery–neck interface. However, the surfaces of the coils farther away than this region harbored no endothelial cells. Scattered inflammatory cells, including neutrophils and monocytes, were seen on the coil surface at the neck central area, where the coil surface was bare at the 4 and 8 weeks’ follow-up.
Conclusions Platinum coil embolization supports gradual but limited endothelialization, where endothelial cells migrate directly from the adjacent parent artery.
- Aneurysm
- Vessel Wall
Statistics from Altmetric.com
Introduction
The precise cellular response to platinum microcoil embolization of saccular aneurysms remains unclear. Rapid aneurysm thrombosis probably relates to coil-induced stasis, but subacute and chronic healing patterns have been poorly characterized to date.1–3 The final common pathway of full endothelialization represents an ideal outcome, allowing permanent closure of the treated aneurysm.4–6 Human histologic samples have shown, even long after embolization, that varying degrees of endothelialization are present.5 ,7 Animal studies have shown variation in the degree of endothelialization, and have also reached disparate conclusions about the exact source of endothelial cells over the neck.8–10 Specifically, the relative importance of circulating versus local cells as precursors to endothelial cells over the treated aneurysm neck has yet to be determined; such a determination would facilitate rational design of future devices. In this study, we used whole tissue mounting staining for endothelial cells on coiled aneurysms to assess the extent and pattern of endothelial cells at the neck following the platinum embolization.
Materials and methods
In vivo experiments
The Institutional Animal Care and Use Committee approved all procedures before the start of the study. Twelve elastase-induced saccular aneurysms were created in female New Zealand white rabbits (2.5–4 kg).11 Aneurysms were permitted to mature for at least 21 days after creation. Animals were anesthetized and a cutdown was performed to gain access to the right common femoral artery. A microcatheter was inserted coaxially through the guiding catheter into the aortic arch. Heparin (100 U/kg) was administered intravenously. The size of the aneurysm cavity was assessed by direct comparison with radiopaque sizing devices during digital subtraction angiography (DSA). The volumetric occlusion rate was calculated as follows: volume=π (dome)2×(length)/6. Considering the aneurysm as a two-dimensional ellipsoid, we assumed the value of the aneurysm dome in this formula was equal to that of the aneurysm width. Volumetric occlusion was calculated in real time, during aneurysm embolization, using the AngioCalc tool (http://www.angiocalc.com). Appropriate sized platinum coils (Heraeus Precious Metals GmbH and Co, Germany; diameter 0.0008 inches) were placed into the aneurysm, as in typical practice. Before and after the placement of each successive coil, volumetric occlusion was calculated. Aneurysm cavities were packed as tightly as possible (>30% packing density) in all cases. Final DSAs were performed immediately after embolization.
Animals were given sustained release (SR) buprenorphine subcutaneously once after coil embolization and were not treated with antiplatelet agents. Four weeks (n=6) and 8 weeks (n=6) after embolization, the coiled aneurysms were harvested for further study.
Tissue harvest and processing
Follow-up DSA was performed, and the animals were then euthanized by a lethal injection of pentobarbital. Median sternotomy and pericardiotomy were performed. Access to the left ventricle was obtained by direct puncture with a 20 G catheter. A small cut was made at the right atrial appendage. By means of a pressure pump, heparinized saline (100 U/mL) was continuously infused into the catheter, until the effluent from the right appendage was light pink. The animals were then perfusion-fixed through the 20 G catheter with 10% buffered formalin for 10 min followed by flushing with heparinized saline for 5 min. The coiled aneurysm was then harvested and immersed in Tris-buffered saline (TBS). Under a dissection microscope (Leica MZ 125), the parent artery was cut longitudinally to expose the neck orifice for gross inspection to evaluate the tissue growth at the neck; the sample was then photographed using the MicroPublisher 5.0 RTV camera attached to the dissection microscope. After photography, the sample was fixed in 10% formalin for 2 h for further whole tissue mount staining. The adjacent left common carotid artery was harvested for positive and negative control.
Whole-mount en face immunostaining
The freshly harvested coiled aneurysms were fixed in 10% neutral buffered formalin for 2 h at room temperature (RT), then washed with TBS for 15 min with four changes. Next, the aneurysms were incubated with 5% normal donkey serum in TBS and Tween-20 for 1 h at RT, followed by overnight incubation with primary antibodies against CD31 (1:30; Dako, Carpentaria, California, USA), or smooth muscle actin (SMA) (1:20, Dako) at 4°C. The aneurysms were washed in TBS for 15 min with three changes, then incubated with a secondary antibody (Cy3-conjugated donkey anti-mouse IgG, 1:200; Jackson Immuno Research, West Groove, Pennsylvania, USA) for 2.5 h at RT, and finally, washed in TBS for 15 min with four changes. Sytox green (1:1000; Life Technologies, Grand Island, New York, New York, USA) served as a nuclear counterstain to identify inflammatory cells. Finally, samples were viewed and imaged with a fluorescence confocal microscope. A positive control was performed using the left common carotid artery with the same antibodies; a negative control was performed with non-immune normal serum in place of the primary antibody.
Results
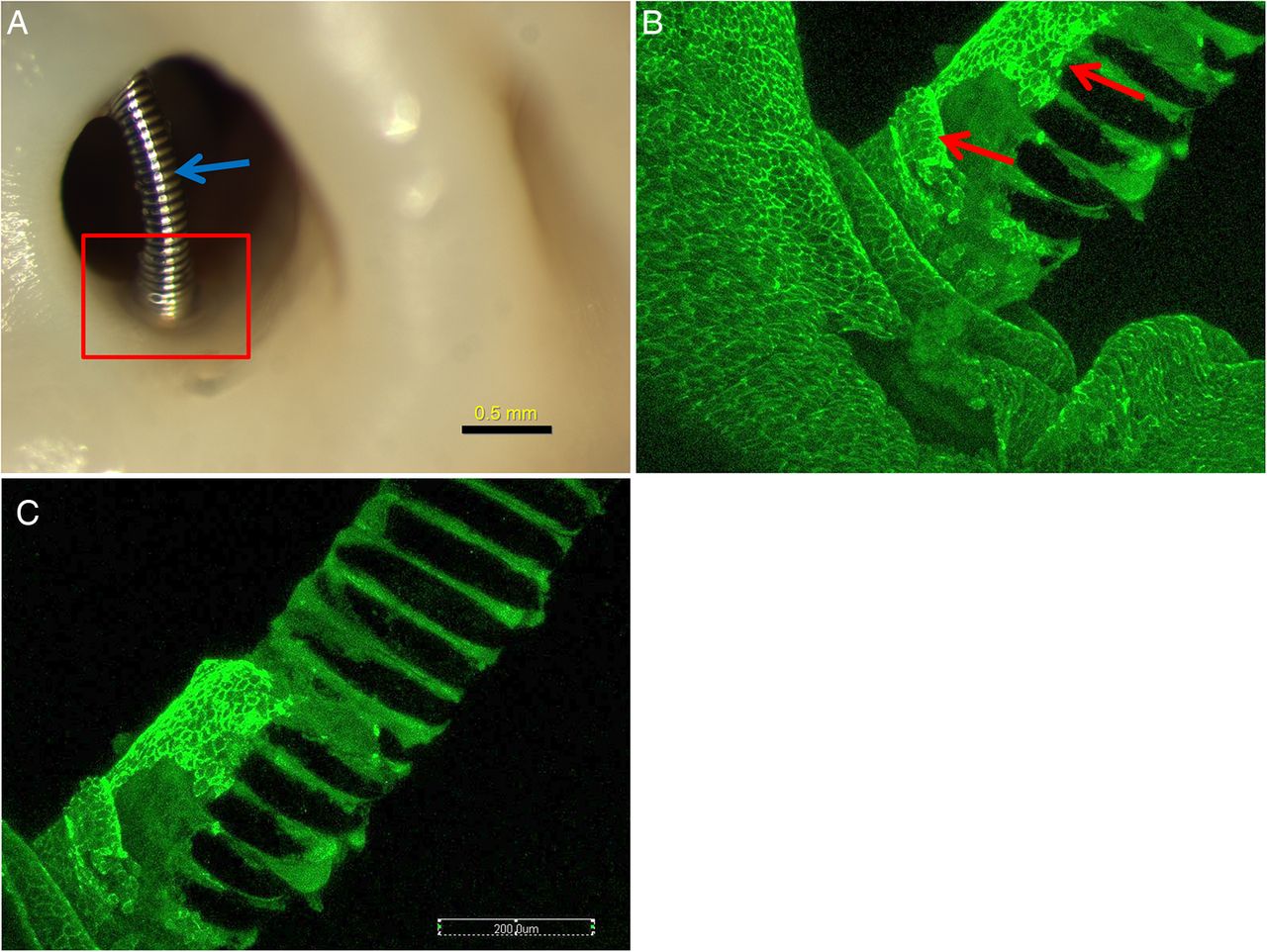
All 12 rabbits survived for the assigned time points and were submitted for histological evaluation. At 4 weeks, two (33%) of six aneurysms did not have coil loops over the neck orifice, with all the coil loops sitting deep within the aneurysm cavity, making dual immunostaining difficult. Four (67%) of six aneurysms had coil loops crossing over the neck orifice and touching the parent artery wall. In the peripheral portions of the neck, limited, white, thin membranous tissue covered the coil loops; over the central areas of the neck the coil loops were bare, without overlying tissue (figure 1A). The en face whole tissue mounting staining showed very localized, single layers of CD31+ endothelial cells corresponding to the thin, membranous areas noted above; these endothelial cells were present at the periphery only, were contiguous with the cells along the parent artery, and extended no more than 400 µm from the parent artery–neck interface (figure 1B, C). In the central areas, where the coil loops extended over the neck without touching the wall, neither CD31+ or SMA+ cells were detected; a few scattered inflammatory cells were present on the coil surface. Scattered and diffused inflammatory cells, including neutrophils and monocytes, were seen on the coil surface or between the coil loops; neither CD31+ nor SMA+ cells were detected on or between the coil loops.

Aneurysm harvested at 4 weeks after embolization. (A) Macrophotograh showing the localized membrane covering at the neck peripheral area (red rectangle), the coil segment which is far away from the wall (blue arrow) and is bare, without tissue coverage. (B) and (C) Whole tissue mount staining (antibody for CD31). The photograph is taken from the rectangular area in (A) showing the localized CD31+ cells (red arrows) covering the coil at the peripheral area, and contiguous with the endothelial cells of the parent artery wall. The portion of the coil which is far away is bare, without CD31+ cell coverage.
At 8 weeks, one aneurysm had coil loops deep within the cavity, which made confocal analysis impossible; this subject was excluded from further analysis. In the remaining five rabbits, four (80%) of five aneurysms had coil loops crossing over the neck orifice and touched the parent wall peripherally. Where the coil loop touched the parent artery wall, localized, thin membranous tissue was grossly visible (figure 2A) and confluent CD31+ endothelial cell covering was detected (figure 2B). Compared with the aneurysms in the 4 week group, the endothelial cells in the 8 week group were more confluent, and extended along the coil loops towards the central area (figure 3C, D), rather than localizing at the periphery. Furthermore, membranous tissue lined with CD31+ cells was also present within the gaps between the coil loops at or near the neck central area (figure 3A, B). However, the maximum distance from the parent artery–neck interface to the leading edge of the endothelialization did not exceed 900 µm in any subject. On the coil loop surfaces, where there was CD31 staining, a few sparse SMA+ cells were present, with some covered with CD31+ cells (figure 2E). Scattered inflammatory cells, primarily monocytes, were present on the coil surface where the coil loops were bare, without CD31+ cell coverage.

Aneurysm harvested at 8 weeks after embolization; (A) Macrophotograph showing the coil loops at the neck orifice. Very thin membrane covering the coil loops at the peripheral area can be seen (red circle). (B) Confocal microphotograph taken from the red circle area in (A) showing that the coil loops at the peripheral area in (A) within the red circle are completely covered with CD31+ cells (large red circle area); those cells are confluent and contiguous with the endothelial cells of the parent artery wall. (C) and (D) are taken from the rectangular area in (A), showing CD31+ cells covering the coil surface; these cells are not limited to the peripheral area but extend to the center. (E) Double staining for CD31 (green) and smooth muscle actin (SMA) (red), showing SMA-positive cells together with CD31+ cells on the coil surface at the peripheral area (whole tissue mount staining: antibody for CD31 (A–D); double staining for CD31 and SMA (E) original magnificent ×20, water lens).

{kind=link}
{kind=link}
{kind=link}
Aneurysm sample harvested at 8 weeks after embolization. (A) Macrophotograph showing the membrane tissue between two coil segments at the central area (red arrow). (B) Confocal microphotograph showing the CD31+ cell lining between the coil segments at the central area shown in (A) (red arrow) (whole tissue mount staining for antibody CD31, original magnification, ×20 water lens).
Discussion
In this study, we showed that bare platinum coils can induce endothelial cell growth. However, at least up to 2 months after embolization, such growth is quite limited and is dependent on close contact between the coils and the junction of the neck and the parent artery. These findings strongly suggest that donor endothelial cells are resident along the parent artery rather than coming from circulating sources. These findings are important as they provide baseline control data for future studies assessing modified coils, and also provide guidance to investigators aiming at improving endothelialization over microcoils used to treat saccular aneurysms.
Endothelialization starts as early as 7 days after treatment in rabbit and swine aneurysm models following embolization.12 ,13 To examine progression of the endothelialization, we chose 4- and 8-week time points for the en face endothelial staining. Both bone marrow-derived circulating stem cells and migration of endothelial cells from the adjacent vessel have been reported in the endothelialization of the aneurysm neck after treatment with flow diverters.12 ,14 Studies have shown that endothelial progenitor cells can promote neck endothelium formation and help the aneurysm to repair post-coil embolization.15 ,16 However, endothelial denudation has been found to prevent recanalization after coil embolization in a canine vein pouch bifurcation model.17 Pandey et al18 developed an in vitro model for studying the mechanism of endothelial cell differentiation and proliferation on endovascular coils by seeding human umbilical vein endothelial cells with coil segments and found that endothelial cells attached to the coils within hours of incubation. They also observed expression of integrins, which promote cell adherence to the extracellular matrix, in the coils seeded with endothelial cells and reported that cell cycle and cell signaling pathways in endothelial cells were affected by platinum coils.
Hemodynamic factors may also influence the growth of endothelial cells at the aneurysm neck, depending on the aneurysm model, and may influence the rate of endothelialization. The alteration of hemodynamics in the aneurysm after coil embolization is believed to promote adhesion of thrombocytes to the coil loops followed by fibrous organization and endothelialization.19–21 In addition, the shape of the coil and configuration might also affect the endothelialization process. Reinges et al8 compared endothelialization of aneurysms embolized with bare, bioactive and hydrogel-coated coils in the rabbit model and suggested that a smooth and dense surface over the aneurysm neck is important for endothelial cells to bridge the aneurysm neck. The lack of neointimal coverage at the aneurysm neck leads to coil compaction and subsequently, recanalization of the aneurysm.22 The growth of neointima derived from the adjacent wall seen in this study is similar to the findings noted in aneurysms treated with a flow diverter.12 Strategies that could promote endothelialization and neointimal growth at the neck would accelerate the permanent healing of aneurysms.
There are several limitations to our study. First, a 2-month follow-up might not have been long enough to monitor the complete growth pattern of endothelial cells following coil embolization. Second, although the results of this study suggested that the neoendothelial cells over the coil loop at the neck were originally from the wall of the parent artery, the exact mechanism of neoendothelial cell proliferation or migration after coil implantation remains unclear. Third, the assessment of endothelialization is only qualitative and the confocal microscopy is limited to evaluation of the coils at the level of the neck.
Conclusion
A platinum coil can induce growth of endothelial cells, which starts primarily at the periphery where the coil loop touches the parent wall. The neoendothelial cells at the aneurysm neck following endovascular device embolization probably originate from the parent artery wall.
References
Footnotes
Contributors Y-HD and IR contributed to the animal experiments and analysis of angiographic data. DD contributed to the tissue staining, interpretation of data and drafting of the manuscript. DK and RK contributed to the conception and design and revised the article critically for important intellectual content.
Funding This work was supported by National Institutes of Health grant NS 076491.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement All authors have access to the raw data.